
0371-60972711
來源: 點擊:1305 喜歡:0
2017-05-12 15:40:34 來源: 點擊:
抗生素自19世紀末被發現以來,廣泛應用于人類感染性疾病的控制和畜禽養殖、水產養殖.但抗生素的不合理使用和長期濫用使得環境中微生物耐藥性產生的選擇壓力增強,導致環境中抗生素抗性細菌 (antibiotic resistance bacteria,ARB) 和抗性基因 (antibiotic resistance genes,ARGs) 含量不斷增加,且抗性基因能通過遺傳和水平基因轉移在親代和其他菌種間傳播擴散,對人類健康和生態平衡造成危害.抗生素和抗生素抗性基因已經被認為是新型環境污染物.污水處理廠是人類活動產生污水的交匯點,同樣也是抗性細菌和抗性基因的儲存庫.目前的研究表明,現有的各類污水處理廠工藝對抗性基因有一定的去除效果,但是出水中抗性基因仍處于一個較高的豐度,存在一定的環境風險.故需要有針對性地研究抗性基因污染問題,結合現有的污水處理工藝,提出合適的控制技術.消毒是污水處理過程中殺滅微生物的水處理過程,是保障污水處理廠尾水水質安全必不可少的環節.消毒處理在殺滅微生物的同時,也可能削減抗性基因.但是,目前對不同消毒削減抗性基因的影響因素和具體機制仍尚不明確.此外,消毒對抗性基因的削減效果還受到其抗性細菌影響,不同抗性細菌對其攜帶的抗性基因可能有不同的保護作用. Huang等對1株未攜帶抗性基因的大腸桿菌和攜帶tetA的大腸桿菌進行紫外和氯消毒研究發現,攜帶抗性基因的菌株表現出更高的消毒耐受性.雖然目前對于氯消毒、紫外消毒、臭氧消毒等常規消毒方法殺滅細菌的機制有了較為成熟的理論,但抗性基因在消毒過程中的行為特征有待討論.本研究采用“消毒+DNaseI酶”實驗來研究抗性基因在各消毒過程中的行為特征.有研究表明,DNA可游離于細胞體之外,成為游離態DNA,并在環境中維持較長時間.游離態DNA上的抗性基因則可能通過基因水平轉移途徑重新進入細菌,繼續產生環境危害. DNaseI酶是1種DNA降解酶,應用其對水環境中胞外游離態DNA進行降解,有助于了解消毒方法削減水中抗性基因的機制.
磺胺類藥是人工合成的抗生素,自1935年臨床應用以來,僅80余年就已產生明顯的環境耐藥性問題.本研究從磺胺類抗性基因入手,分析氯消毒、紫外消毒、臭氧消毒等常規消毒方法對抗性基因的削減作用.從某污水處理廠二級出水中分離出磺胺類抗性細菌,并挑取典型的革蘭氏陽性菌和陰性菌各1株,確定其中的磺胺類抗性基因種類及菌株種類.以上述兩株細菌為實驗對象,污水廠二級出水滅菌后作為實驗水樣,控制上述細菌濃度投加到實驗水樣中,比較研究消毒對抗性細菌和抗性基因的削減效果.同時,利用“消毒+DNaseI酶”實驗研究磺胺類抗性基因在消毒過程中的行為特征,以期為理解消毒削減抗性基因的機制,并為污水中抗性基因污染的控制方法提供新思路.
1 材料與方法1.1 實驗菌株篩選
采集浙江臨安某污水處理廠氧化溝出水 (二級出水),采用稀釋、涂布分離方法在磺胺類選擇培養基平板上篩選磺胺類抗性細菌.磺胺類選擇培養基配置方法為:蛋白胨10 g ·L-1,酵母提取物5 g ·L-1,氯化鈉10 g ·L-1,調節pH至7后121℃、20 min高壓滅菌,在完全凝固前趁溫加入磺胺甲唑母液至濃度為50.4 μg ·mL-1.母液濃度40 mg ·mL-1,采用1 mol ·L-1 NaOH溶液在避光條件下溶解配置. 30℃、24 h培養后,挑取單菌落分離純化,采用細菌基因組提取試劑盒TIANamp Bacteria DNA Kit (TIANGEN, China) 提取細菌DNA,由上海睿迪生物科技有限公司測定16S rRNA序列,測序結果通過使用NCBI網站 (http://www.ncbi.nhn.nih.ov/blast/) 的BLAST進行序列同源性檢索比對,確定單菌株菌種.選擇典型革蘭氏陽性菌和革蘭氏陰性菌各1株,采用普通PCR法對兩株細菌分別進行磺胺類抗性基因 (sul Ⅰ 、sul Ⅱ 、sul Ⅲ 、sulA) 及16S rRNA、intI1基因進行定性檢測,分析其基因類型.
PCR體系為:10×PCR buffer (Mg2+)2.5 μL,2.5 mmol ·L-1,dNTP 2.0 μL,10 mmol ·L-1上游引物1.0 μL,10 mmol ·L-1下游引物1.0 μL,5U Taq酶0.3 μL,ddH2O 17.2 μL,DNA樣品1.0 μL.各基因引物見表 1.在PCR循環儀 (TIANLONG,China) 中進行擴增,PCR擴增條件為:95℃預變性5 min后,95℃變性15 s,退火30 s,72℃延伸30 s,共35個循環,最后72℃下延伸7 min.普通PCR完成后,取5 μL PCR反應產物,與6×Loading buffer Dye混合均勻后,使用經EB染色的1.5%的瓊脂糖凝膠,在120 V電壓下電泳30 min檢測擴增條帶.使用全自動凝膠成像儀 (PEIQING, China) 對瓊脂糖凝膠進行拍照,分析所得PCR產物長度并與表 1中提供的擴增片段相比較.
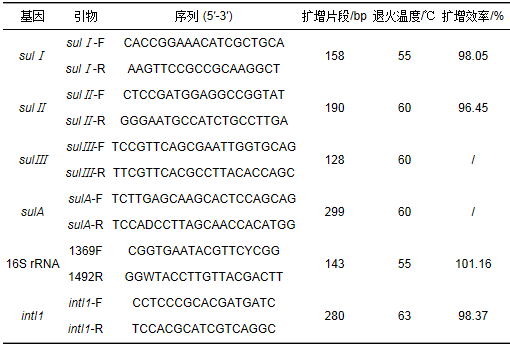
表 1 各基因引物序列及退火溫度
本研究采用菌株為:①瓊氏不動桿菌,Acinetobacter junii,革蘭氏陰性,16S rRNA序列NCBI登錄號:KY120958;②枯草芽孢桿菌,Bacillus subtilis,革蘭氏陽性,16S rRNA序列NCBI登錄號:KY121111.兩者均含有sul Ⅰ、sul Ⅱ ,不含有sul Ⅲ、sulA,故下文對磺胺類抗性基因的研究主要集中于sul Ⅰ、sul Ⅱ .
1.2 實驗水樣準備
將實驗菌株培養30℃、24 h后的菌液10 000 r ·min-1離心10 min,棄上清液收集菌體,用無菌PBS重懸,利用0.5號麥氏比濁管控制細菌濃度約為1.5×108 CFU ·mL-1.污水處理廠二級出水采用0.22 μm膜進行過濾滅菌后,將細菌PBS溶液以1 :14的比例投加至水樣中,使最終實驗水樣細菌濃度約為107 CFU ·mL-1(模擬現實水樣細菌濃度).
1.3 氯消毒
氯消毒劑采用次氯酸鈉溶液,將其稀釋至有效氯10 g ·L-1,并置于棕色試劑瓶中在4℃下避光保存,實驗前測定實際有效氯含量后立即使用.取1 L水樣,滴加次氯酸鈉溶液,使樣品中的有效氯濃度依次達到設定值 (1、2、3、4、5 mg ·L-1).實驗水樣體積為1 L,氯消毒接觸時間為30 min.在氯消毒完成后,投加5 mg ·L-1的硫代硫酸鈉溶液終止消毒反應.每次消毒實驗重復3次,下同.
1.4 紫外消毒
紫外線消毒處理在實驗室自主設計的紫外線平行光束儀裝置內進行,平行光束儀內含有5根不同功率的低壓紫外UV254燈管,燈光水平中心線距離實驗水樣表面50 cm.紫外線平行光束儀裝置示意如圖 1所示,從燈管發出的紫外線經平行光柱,均勻地輻照到反應容器 (20 cm×20 cm×5 cm).實驗水樣體積為1 L,控制紫外燈強度不變,通過調節照射時間以獲得不同的紫外劑量 (5、10、15、20、25 mJ ·cm-2).為避免光復活效應,經紫外消毒處理后,立即進行下一步的樣品處理和檢測.

圖 1 紫外消毒與臭氧消毒裝置示意
1.5 臭氧消毒
臭氧消毒處理采用VMUS-DG臭氧發生器 (AZCO,Canada).氣路上裝有流量計和臭氧分析儀UV100(ECO,USA),用以測量進出氣體中的臭氧濃度,如圖 1所示.實驗水樣體積為1 L,通過調節氣體流速和臭氧濃度,使臭氧消毒接觸時間為10 min時,臭氧投加量達到目標設定值 (2、4、6、8、10 mg ·L-1).
1.6 DNaseI處理
向消毒處理后的水樣中加入500 U單位DNaseI (約可降解250 μg DNA,大于水樣中總DNA含量),15℃反應振蕩2 h.
1.7 菌落計數法
本研究使用菌落計數法來確定水樣中的磺胺類抗性細菌數目.用無菌水對水樣進行梯度稀釋,各取100 μL水樣稀釋液于磺胺類選擇培養基平板上,用玻璃刮刀涂布均勻,平行3份,30℃、24 h培養后,進行菌落計數 (CFU ·mL-1).
1.8 熒光定量PCR
本研究使用熒光定量PCR法檢測抗性基因的絕對豐度.水樣采用FastDNA Spin Kit forsoil試劑盒 (MP,USA) 提取DNA,使用微量蛋白質核酸分析儀 (Nanodrop,USA) 測定DNA濃度,置于-85℃保存.
熒光定量PCR標準曲線的建立:普通PCR擴增得到的目的基因序列使用Biospin凝膠回收試劑盒 (BIOER,China) 進行純化回收PCR產物,后測量DNA含量、純度并調節至合適濃度后,連接pMD19-T載體,轉化入E. coli感受態細胞DH5α,將感受態細胞涂于含有氨芐青霉素、X-gal和IPTG的LB固體培養基上培養12~16 h;通過藍白斑篩選挑取白色重組菌斑,挑選陽性克隆子用LB培養液擴大培養,待菌液混濁后,取1 mL菌液測序插入基因片斷.測序結果通過使用BLAST進行序列同源性檢索比對;其余3~4 mL菌液用來提取質粒,使用QIAGEN質粒專用提取試劑盒 (QIAPREP,Germany) 進行質粒抽提,確保質粒DNA的A260/A280比值在1.8左右.符合要求的質粒,作為標準品計算其濃度[質粒濃度的計算公式為:濃度=(質量÷相對分子質量)×6.02×1023].將已知拷貝數的質粒標準品依次進行10倍稀釋,并且保持質粒濃度在108~102之間.標線擴增效率見表 1.
定量PCR反應在Bio-rad IQ5熒光定量PCR儀器 (Bio-Rad,USA) 中進行.定量PCR反應的反應液體系:7.5 μL SYBR Premix Ex Taq溶液,0.3 μL ROX Reference溶液,0.3 μL濃度為10 mmol ·L-1濃度的正向引物,0.3 μL濃度為10 mmol ·L-1濃度的反向引物,4.6 μL ddH2O.反應液體系在100 μL 96孔反應板的反應孔中進行.定量PCR反應程序為預變性95℃下熱變性30 s;然后進入40個循環的擴增階段,包括95℃變性5 s,退火30 s,72℃延伸30 s,延伸的同時掃描熒光信號.溶解曲線程序為55~95℃之間,每0.5℃讀數,其間停留30 s.每個樣品做3次重復.
1.9 數據處理
數據采用Excel 2010和Origin 9.0軟件進行分析.
2 結果與討論2.1 3種消毒方法對磺胺類抗性細菌的削減效果
不同消毒方法對實驗水樣中的磺胺類抗性菌株均有一定的去除效果,且在低劑量濃度下,抗性細菌的削減效果隨著消毒劑量的增加而提高,在實驗設置的最高劑量 (5 mg ·L-1氯投加濃度,25 mJ ·cm-2紫外劑量,10 mg ·L-1臭氧投加濃度) 下,實驗水樣中的抗性細菌大多已被滅活,如圖 2所示.當消毒劑量達到一定水平后,抗性細菌均不能被菌落計數法檢測.在兩種細菌的比較中,實驗發現枯草芽孢桿菌 (革蘭氏陽性菌) 的耐受性強于瓊氏不動桿菌 (革蘭氏陰性菌),這可能與革蘭氏陽性菌的細胞壁保護作用有關,其能阻礙自由氯和臭氧分子的穿透,一定程度上保護細菌.在紫外消毒中,McKinney等[10]也發現了類似現象,他們深入研究后認為,這可能與兩種細菌基因序列中總基因組大小不同有關.紫外消毒導致DNA中的嘧啶經過光化學的破壞,相鄰胸腺嘧啶產生二聚作用.細菌的DNA中多數胸腺嘧啶二聚物的形成阻止了DNA復制和最終導致細胞死亡,達到消毒的目的.革蘭氏陽性菌的總基因組更小,意味著它們有更少的潛在嘧啶二聚體目標,所以枯草芽孢桿菌對紫外的敏感性更低.

G-ARB:瓊氏不動桿菌水樣;G+ARB:枯草芽孢桿菌水樣;ARB存活率為存活量與初始量的比值,兩者單位為CFU ·mL-1
圖 2 不同消毒方法中磺胺類抗性細菌數量的變化
對消毒劑消毒效果的比較,可以采用CT值 (concentration-time value) 來衡量. CT值是消毒劑的濃度和作用時間的乘積,用于比較消毒劑殺菌作用的指標,CT值越低消毒效果越好.將氯消毒和臭氧消毒相對比,當瓊氏不動桿菌的削減率達到3個數量級左右時,氯消毒的CT值為90 mg ·(min ·L)-1,而臭氧消毒的CT值為20 mg ·(min ·L)-1,可以看出,臭氧的CT值遠遠小于氯.因此臭氧削減目標細菌所需時間更短,效率更高.此外,污水廠常使用紫外消毒作為深度處理工藝,使用的輻照劑量在30 mJ ·cm-2以下.本研究中,25 mJ ·cm-2消毒劑量下,瓊氏不動桿菌和枯草芽孢桿菌基本滅活,研究表明通常污水廠所設置的輻射劑量能滅活磺胺類抗性細菌.
2.2 3種消毒方法對磺胺類抗性基因的削減效果
目前各消毒方法的評價都是以微生物的削減程度來衡量的,但是根據實驗結果,發現削減微生物的量并不直接等同于削減抗性基因的量. 圖 3為實驗水樣在不同消毒劑量下磺胺類抗性基因的變化情況,可知在消毒過程中,磺胺類抗性基因大體上是隨著消毒劑量的增加而下降的.氯消毒中削減效果較為明顯,在本實驗設置的最低濃度1 mg ·L-1時,瓊氏不動桿菌水樣的sul Ⅱ 絕對豐度削減率為78.9%,枯草芽孢桿菌水樣的sul Ⅱ 絕對豐度削減率為70.5%. sul Ⅱ 的削減率要高于sul Ⅰ ,但是隨著消毒劑量的上升,sul Ⅰ 基因豐度也有明顯的下降,氯消毒濃度達到3 mg ·L-1,枯草芽孢桿菌水樣的sul Ⅰ 絕對豐度削減率由2 mg ·L-1的29.7%上升到77.9%.在氯消毒濃度達到4 mg ·L-1時,兩種不同目標細菌sul Ⅰ 、sul Ⅱ 絕對豐度削減率都能達到95.0%以上.瓊氏不動桿菌水樣與枯草芽孢桿菌水樣磺胺類抗性基因的基因豐度隨氯消毒濃度的變化規律基本一致,但瓊氏不動桿菌水樣的削減率高于枯草芽孢桿菌水樣.結合磺胺類抗性基因豐度和16S rRNA基因豐度之間的相關性分析發現,氯消毒過程中,瓊氏不動桿菌水樣中sul Ⅱ 基因與16S rRNA基因呈現正相關關系 (P < 0.05),R2為0.90(如圖 4);枯草芽孢桿菌水樣中sul Ⅰ 、sul Ⅱ 基因均與16S rRNA基因呈現正相關關系 (P < 0.05),R2分別為0.77和0.59. 16S rRNA基因的豐度在一定程度上反映了樣品中的微生物量,這表明氯消毒削減磺胺類抗性基因主要和微生物量的降低有關.

G-sulⅠ、G-sulⅡ:瓊氏不動桿菌水樣中sulⅠ、sulⅡ基因;
G+sulⅠ、G+sulⅡ:枯草芽孢桿菌水樣中sulⅠ、sulⅡ基因
圖 3 不同消毒方法中磺胺類抗性基因絕對豐度的變化

圖 4 氯消毒中瓊氏不動桿菌16S rRNA與sulⅡ擬合曲線
紫外消毒和臭氧消毒對磺胺類抗性基因的削減效果一般,在設置的最高劑量 (25 mJ ·cm-2紫外劑量,10 mg ·L-1臭氧投加濃度) 下,sul Ⅰ 絕對豐度削減率為84.9%、84.6%(瓊氏不動桿菌) 和77.4%、84.3%(枯草芽孢桿菌),sul Ⅱ 絕對豐度削減率為89.9%、86.2%(瓊氏不動桿菌) 和85.7%、87.2%(枯草芽孢桿菌),抗性基因絕對豐度僅下降不到一個數量級,仍處于較高程度.同時,有研究認為,低劑量氯消毒 ( < 40 mg ·min ·L-1) 促進細菌接合轉移,在現實環境中,消毒對抗性基因的抑制作用可能更弱.臭氧消毒過程中磺胺類抗性基因的削減規律呈現波動,臭氧消毒濃度為0~6 mg ·L-1時,削減情況呈現波動,對磺胺類抗性基因基本無削減效果 (平均削減率為9.1%),濃度達到8 mg ·L-1時平均削減率升至55.3%,呈現一定下降趨勢,可以認為低濃度臭氧消毒對磺胺類抗性基因的削減效果并不明顯.比較臭氧消毒和氯消毒對磺胺類抗性基因的削減效果,當枯草芽孢桿菌sul Ⅰ 絕對豐度削減率達到80.0%左右時,氯消毒的CT值為90 mg ·(min ·L)-1,而臭氧消毒的CT值為100 mg ·(min ·L)-1,其他情況下也能分析到相似結論,這與削減抗性細菌的結果截然相反.氯消毒過程中,抗性基因的削減與微生物量減少相關,這表明臭氧消毒中,抗性基因的減少可能不是由于微生物量的減少,抗性細菌減少時抗性基因仍得到一定程度的保存.同時,磺胺類抗性基因豐度和16S rRNA基因豐度的相關性分析表明,兩種水樣sul Ⅰ 、sul Ⅱ 基因均與16S rRNA基因相關性不顯著 (P>0.05).臭氧消毒和紫外消毒中,抗性細菌被殺滅后,抗性基因的行為特征需要進行深入研究.
此外,對兩種細菌進行對比發現,兩者在氯消毒和紫外消毒過程中的變化規律基本一致,但是瓊氏不動桿菌 (革蘭氏陰性菌) 中磺胺類抗性基因的削減率高于枯草芽孢桿菌 (革蘭氏陽性菌).在3 mg ·L-1氯消毒濃度時,瓊氏不動桿菌sul Ⅰ 、sul Ⅱ 基因削減率分別為94.8%和97.4%,而枯草芽孢桿菌sul Ⅰ 、sul Ⅱ 基因削減率為77.8%和83.3%,表明磺胺類抗性基因在不同種類細菌中對氯消毒的耐受性有一定差別,革蘭氏陽性菌耐受性強于革蘭氏陰性菌,這與抗性細菌的削減情況相同,符合氯消毒削減磺胺類抗性基因主要和微生物量的降低有關的結論.在10 mJ ·cm-2紫外消毒劑量時,瓊氏不動桿菌sul Ⅰ 、sul Ⅱ 基因削減率分別為71.4%和60.0%,而枯草芽孢桿菌sul Ⅰ 、sul Ⅱ 基因削減率為37.2%和38.6%,表明紫外消毒中抗性基因的削減仍受到細菌種類的影響.值得注意的是,有研究對污水紫外消毒進行深入研究發現,UV消毒在減小細菌多樣性的同時,會提高抗性基因在污水中的相對豐度.抗生素抗性細菌和非抗性細菌在紫外消毒過程中的不同表現同樣會減少抗性基因的削減率.在臭氧消毒中,兩種細菌的削減率變化無明顯規律,臭氧消毒可能與細菌種類無明顯關系.
2.3 應用DNaseI研究消毒中磺胺類抗性基因削減的機制
3種消毒方法均能削減兩種目標細菌豐度和抗性基因豐度,但細菌和基因豐度的削減程度相差很多,例如20 mJ ·cm-2紫外消毒下,消毒前后細菌濃度相差5個數量級,而抗性基因的豐度相差僅為0~1個數量級,磺胺類抗性基因豐度仍高達105~106 copies ·mL-1.表明基因在其細菌被殺死后可能仍獨立于細胞體外存留,抗生素抗性基因亦可能包含其中,并在特定條件下,又會進入其他細胞中重新表現出耐藥性,從而繼續傳播.本研究選擇3 mg ·L-1(氯消毒)、10 mJ ·cm-2(紫外消毒)、6 mg ·L-1(臭氧消毒) 劑量消毒后實驗水樣,進行DNaseI處理,圖 5為DNaseI處理前后實驗水樣中磺胺類抗性基因豐度的變化情況.經過DNaseI處理后,目標細菌磺胺類抗性基因豐度均有明顯的下降.其中,臭氧消毒前后磺胺類抗性基因豐度變化最大,例如瓊氏不動桿菌氯消毒、紫外消毒水樣中sul Ⅰ 基因在DNaseI處理前后比例分別為28.5%和67.5%,而臭氧消毒水樣在DNaseI處理后僅剩4.3%,即有95.7%的去除率,明顯高于氯消毒和紫外消毒.氯消毒水樣中經過DNaseI處理后的去除率基本高于紫外消毒.實驗表明,臭氧消毒后,磺胺類抗性基因主要存在于水環境中的游離態DNA.

圖 5 DNaseⅠ處理前后磺胺類抗性基因豐度的變化
對DNaseI處理前后16S rRNA和intⅠ1基因變化分析發現 (見圖 6),DNaseI對16S rRNA基因豐度有一定的削減效果 (14.6%~74.0%),總體削減率在50.0%左右.其中在紫外消毒和氯消毒后DNaseI對16S rRNA基因豐度的削減率在42.0%~74.0%,削減率相對于臭氧消毒高.在不同消毒方法處理后DNaseI對intⅠ1基因豐度的削減情況不相同,臭氧消毒后DNaseI對intⅠ1基因豐度削減率最高,氯消毒和紫外消毒后DNaseI對intⅠ1基因豐度削減率低. intⅠ1基因作為1種水平基因轉移元件,可以認為是抗性基因在游離態DNA中的主要載體之一.結合上文研究,氯消毒過程中磺胺類抗性基因與16S rRNA的豐度顯著相關,而游離態DNA中磺胺類抗性基因的削減情況基本與16S rRNA的削減情況一致,這均表明,氯消毒對磺胺類抗性基因的削減主要與減少細菌量有關.在紫外消毒方面,游離態DNA中磺胺類抗性基因的含量不高,而紫外消毒的機制主要是細胞的DNA中的胸腺嘧啶經過光化學的破壞,相鄰的嘧啶產生二聚作用.細菌的DNA中多數胸腺嘧啶二聚物的形成阻止了DNA復制和最終導致細胞死亡,達到消毒的目的.之前的研究結果同樣表明,紫外消毒過程中不同種類細菌所攜帶抗性基因呈現不同的表現,表明紫外消毒對磺胺類抗性基因的削減主要是直接破壞磺胺類抗性基因,達到削減的效果.

G-16S rRNA、G-16S rRNA:瓊氏不動桿菌水樣中16S rRNA基因;G+intⅠ1、G+intⅠ1:枯草芽孢桿菌水樣中intⅠ1基因
圖 6 DNaseI處理前后實驗水樣中16S rRNA和intⅠ1基因
值得關注的是,臭氧消毒中,DNaseI對16S rRNA基因豐度削減率不高 (瓊氏不動桿菌水樣和枯草芽孢桿菌水樣分別為14.6%和25.9%),而對intⅠ1基因豐度的削減相對較高 (瓊氏不動桿菌水樣和枯草芽孢桿菌水樣分別為83.0%和95.5%),這表明游離態DNA中16S rRNA基因含量不高,intⅠ1基因含量高.這與臭氧消毒能有效殺滅目標細菌,而對磺胺類抗性基因削減率不高的實驗現象基本符合.目前對臭氧消毒機制觀點主要有以下兩點:①臭氧能氧化分解細菌內部葡萄糖氧化酶,破壞細胞的核酸,分解DNA、RNA、蛋白質、脂肪和多糖等大分子聚合物,使細菌的物質代謝生長和繁殖過程遭到破壞. ②臭氧能滲透細胞膜組織,侵入細胞膜內作用于外膜脂蛋白和內部的脂多糖,使細胞發生畸變,導致細胞的溶解死亡.這表明臭氧消毒的機制可能主要為侵入細胞膜內作用于外膜脂蛋白和內部的脂多糖,使細胞發生畸變,導致細胞的溶解死亡,大量含有抗性基因的游離DNA就散逸到水環境中,仍具有潛在的環境風險。
3 結論
(1) 氯消毒、紫外消毒、臭氧消毒對城市污水處理廠尾水中磺胺類抗性細菌有較好的滅菌效果,受到消毒劑量和細菌種類的影響,但對磺胺類抗性基因的控制效果不夠理想.
(2) 氯消毒削減磺胺類抗性與細菌量的減少有關,紫外消毒過程中磺胺類抗性基因可能被直接破壞而被削減,而臭氧消毒過程中磺胺類抗性基因隨游離態DNA進入到環境中,存在潛在的環境問題,需要引起關注.
上一篇:含氮廢水生物脫氮工藝研究
下一篇:水處理中超濾工藝研究

